Definition des Hormonbegriffs
Hormone als Signalvermittler
Die Entwicklung vielzelliger Lebewesen aus dem Zusammenschluss von Einzelzellen ist ein ungeheurer Fortschritt in der Evolution gewesen. Eine seiner wesentlichen Voraussetzungen ist die Entwicklung
der Signalübermittlung von Zelle zu Zelle bzw. von Organ zu Organ. Die Signale werden auf humoralem Weg in Form chemischer Verbindungen , nämlich der Hormone ,
als Signalvermittler übertragen.
Erfogt die Signalübermittlung durch Diffusion des hormonellen Faktors von der sezernierenden direkt auf eine benachbarte Zelle , so spricht man von parakriner Sekretion. Einen
Sonderfall stellt die juxtakrine Sekretion dar, bei der der hormonelle Faktor in der Plasmamembran der produzierenden Zelle verankert ist und für die Wechselwirkung mit dem
entsprechenden Rezeptor auf der Zielzelle ein direkter Zell-Zell-Kontakt nötig ist.
Wird dagegen das Hormon einer sezernierenden Zelle in die Blutbahn abgegeben , um seine Funktion an einer weiter entfernten Zelle auszuüben, so handelt es sich um endokrine Sekretion.
Von parakriner und endokriner Sekretion abzugrenzen ist schliesslich die autokrine Sekretion , die als interne bzw. externe autokrine Sekretion vorkommt. Sie beruht darauf, dass
von einer sezernierenden Zelle gebildete Signalmoleküle auf diese Zelle selbst rückwirken.
Grundsätzlich sind Hormone bzw. Zytokine extrazelluläre Signalmoleküle. Damit sie die gewündschte intrazelluläre Antowort auslösen
können, müssen sie mit einem zellulären Rezeptor in Wechselwirkung treten. Derartige Rezeptoren sind immer Proteine, die als integrale Membranproteine , als zytoplasmatische Proteine
oder als Kernproteine vorliegen können. Meist löst der durch die Reaktion von Hormon mit Rezeptor entstandene Hormon-Rezeptorkomplex die Bildung eines intrazellulären Singalmoleküls
aus. Der Mechanismus, mit dem dies geschieht, wird auch als Signaltransduktion bezeichnet. Oft erfolgt die Signaltransduktion in einem mehrstufigen Prozess , was auch als Signalkaskade bezeichnet
wird. Von grosser Bedeutung ist auch die Beendigunng der zellulären Antwort auf einen hormonellen Stimulus. Meist werden dabei die intrazellulär entstandenen Signalmoleküle inaktiviert
. Dieser Vorgang wird auch als Signallöschung bezeichnet.
Glanduläre Hormone, Gewebehormone und Zytokine
Eine Reihe von Hormonen wird in endokrinen Drüsen gebildet und dementsprechend als glanduläre Hormone bezeichnet. Endokrine Drüsen sind
- Die Hypophyse (Vorder- und Hinterlappen)
- Die Langerhans'schen Inseln des Pankreas
- Die Schilddrüse
- Die Nebenschilddrüsen
- Die Nebennieren (Nebennierenmark und rinde)
- Die männlichen und weiblichen Keimdrüsen
- Die Plazenta
Die von diesen Drüsen gebildete Hormone werden ins Blut abgegeben.
Im Gegensatz zu den glandulären Hormonen werden dagegen die Gewebehormone von besonderen, in den verschiedensten Geweben verstreuten, Zellen synthetisiert. Einige Gewebehormone
erreichen ihre Zielzelle ähnlich wie die glandulären Hormone über den Blutweg (z.B. gastrointestinale Hormone, Eikosanoide, Kinine und biogene Amine Histamin, Serotonin). Die Zielzellen
anderer Gewebshormone befinden sich dagegen in unmittelbarer Nachbarschaft zu den hormonproduzierenden Zellen oder sind mit diesen identisch (parakrine bzw. autokrine Sekretion).
Von besonderer Bedeutung innerhalb der Gewebehormone sind die regulatorischen Peptidfaktoren oder Zytokine (EPO, G-CSF, Interferone, Interleukine, insulin-like-growth-factor,
tumor necrosis factor etc.). Sie werden von den verschiedensten Zellen freigesetzt und regulieren meist als parakrine Faktoren Proliferation bzw. Differenzierung und Funktion ihrer Zielzellen.
Einteilung der Hormone und Zytokine
Hormone lassen sich nach funktionellen Aspekten in Gruppen einteilen:
In den ersten beiden Gruppen befinden sich danach diejenigen Hormone, die Wachstums- und Differenzierungsvorgänge beeinflussen.
In der Gruppe I befinden sich die Zytokine , die als Gewebehormone Differenzierung oder Wachstum der unterschiedlichen Zellen beeinflussen (z.B. IGF).
In der Gruppe II befinden sich eine Reihe glandulärer Hormone , die ebenfalls Wachstum und Differenzierung steuern. Neben dem Wachstumshormon gehören
hierzu die Schilddrüsenhormone, Sexualhormone sowie die in der Nebennierenrinde gebildeten Glucocorticoide. Auffallend ist, dass die Aktivität dieser
endokrinen Drüsen durch hypophysäre und hypothalamische Hormone reguliert wird.
In der Gruppe III befinden sich die Hormone, deren Wirkungseintritt sehr rasch, d.h. innerhalb weniger Minuten erfolgt und die aus diesem Grund für die schnelle Umstellung
des Stoffwechsels zu sorgen haben. Zu ihnen gehören v.a. das Insulin , sowie seine Gegenspieler Glucagon und die Katecholamine.
In der Gruppe IV finden sich diejenigen Hormone, die Verdauung und Resorption von Nahrungsstoffen überwachen (gastrointestinale Hormone : Sekretin , Gastrin u.a.)
Die Gruppe V umfasst Hormone, die in den Stoffwechsel von Calcium und Phosphat eingreifen. Es handelt sich um das Parathormon der Nebenschilddrüse, das Thyreocalcitonin der
Schilddrüse sowie die biologisch aktiven Formen des Vitamin D (D-Hormone).
In der Gruppe VI finden sich die Hormone, die den Stoffwechsel von Wasser und Elektrolyten regulieren. Hierzu gehören das Vasopressin aus dem Hypophysenhinterlappen,
das Angiotensin , die Mineralocorticoide der Nebennierenrinde und das natriuretische Atriumpeptid.
Stoffwechsel der Hormone
Biosynthese und Sekretion von Hormonen
Hormone gehören chemisch zu den unterschiedlichsten Verbindungen. Sie können
- Derivate von Aminosäure (Katecholamine, biogene Amine etc.)
- Abkömmlinge des Cholesterins oder mehrfach ungesättigter Fettsäuren
- Peptide und Proteine sein (Insulin, Glucagon
ect)
Dementsprechend unterscheiden sich natürlich auch die Mechanismen ihrer Biosynthese. Besonders die Peptid- und Proteohormone (z.B. Insulin, Glucagon, Parathormon) werden wie andere sekretorische
Proteine in Form höhermolekularer Vorstufen synthetisiert und zum Tiel so gespeichert. Gelegentlich tragen derartige Präkursoren sogar mehrere unterschiedliche Hormone,
die durch entsprechende proteolytische Spaltung freigesetzt werden.
Eine grosse Zahl von Hormonen wird in den endrokrinen Zelle gespeichert. Bei vielen Peptid- und Proteohormonen, aber auch bei Aminosäurederivaten wie den Katecholaminen erfolgt diese Speicherung
in Form intrazellulärer Sekretvesikel. Einen besonderen Fall stellen die Schilddrüsenhormone dar, die als Kolloid extrazellulär in grossen Mengen in der Schilddrüse gespeichert
werden. Andere Hormone , z.B. Steroidhormone oder das PTH, werden nur in geringem Umfang gespeichert.
Transport von Hormonen im Blut
Die glandulären Hormone gelangen über den Blutkreislauf an ihren Wirkungsort. Die vom Cholesterin abgeleiteten Steroidhormone sowie die Hormone der Schilddrüse sind besonders
hydrophob. Aus diesem Grund können sie nur in Bindung an spezifische Transportproteine im Serum transportiert werden.
Abbau und Ausscheidung von Hormonen
Für den Abbau und die Ausscheidung von Hormonen zeichnen sich vor allem die Leber und die Nieren verantwortlich.
Intrazelluläre Hormonrezeptoren
Eine Reihe von Hormonen, so u.a. die Steroid- oder Schilddrüsenhormone, wirken durch Aenderung der Transkription spezifischer Gene. Dies führt zu einer gesteigerten oder gehemmten Biosynthese
eines spezifischen Proteins, meist eines Enzyms, oder einer Gruppe spezifischer Proteine (Enzyme). Der erste Schritt in der Wirkung dieser Hormone ist die Aufnahme in die Zelle und die dortige Bindung
an spezifische Rezeptorproteine, welche cytoplasmatisch oder nukleär lokalisiert sein können.
Die Bindung des Hormons an intrazelluläre Rezeptoren aktiviert diese zu Transkriptions-faktoren. Der Hormonrezeptorkomplex ist ein Transaktivator für spezifische Enhancer-Regionen der
ensprechenden regulierten Gene (Promotor-Regionen). Diese Enhancer (bzw. Silencer) Sequenzen haben meist eine palindromische Struktur.
Membranassozierte Hormonrezeptoren
Viel Hormone zeichnet sich dadurch aus, dass bis zu ihrem Wirkungseintritt nur Sekunden bis Minuten vergehen. Damit ist es wenig wahrscheinlich, dass sie über eine Aenderung der Proteinbiosynthese
wirken. Sie werden von ihren Zielzellen nicht aufgenommen, sondern entfalten ihre Wirkung durch Bindung an einen in der Plasmamembran gelegenen Rezeptor. Dieses Ereignis veranlasst die Bildung
intrazellulärer Botenstoffe. Diese werden auch als second messenger und dementsprechend das Hormon als first messenger bezeichnet.
Liganden-regulierte Ionenkanäle
Hormon- oder allgemeiner Liganden-gesteuerte Ionenkanäle werden durch extrazelluläre Liganden aktiviert. Zu ihnen gehören z.B. die Neurotransmitter Acetylcholin , Glutamat oder Serotonin .
Intrazellulär aktivierte Ionenkanäle spielen eine wichtige Rolle bei der Photorezeption, der Geruchserkennung, aber auch allgemeiner bei der Regulation der intrazellulären Calciumkonzentration
. Liganden-regulierte Ionenkanäle vermitteln die schnellsten bekannten zellulären Reaktionen auf Hormone oder Transmitter , da die Bindung des Liganden unmittelbar
mit der spezifischen Antwort, nämlich dem Oeffnen oder Schliessen eines Ionenkanals verknüpft ist. Anders als bei den anderen Rezeptortypen ist die Erzeugung eines intrazellulären
Boten (second messenger) für die Signaltransduktion nicht notwendig.
Die durch extrazelluläre Liganden aktivierten Ionenkanäle haben eine gemeinsame Grundstruktur. Sie sind jeweils aus fünf Proteinuntereinheiten zusammengesetzt.
Die durch intrazelluläre Liganden regulierten Ionenkanäle sind weniger gut charakterisiert. Aus einer Reihe von Untersuchungen geht jedoch hervor, dass sie häufig aus Untereinheiten
mit sechs Transmembrandomänen bestehen.
Liganden-regulierte Ionenkanäle (Auswahl)

Prinzip der hormonellen Signaltransduktion
Die meisten der in die Plasmamembran integrierten Hormonrezeptoren lösen eine Reihe unterschiedlicher intrazellulärer Antworten aus.
- Sie mobilisieren intrazelluläre Calciumspeicher,
- Sie führen durch covalente Modifikationen zur (In-)Aktivierung von Enzymen
- Sie greifen in die Genexpression ein.
Das generelle Problem bei diesen Vorgängen besteht darin, die extrazelluläre Bindung des Liganden an den Hormonrezeptor in ein intrazelluläres Signal umzusetzen. Intrazelluläre
Signale können dabei kleine Signalmoleküle wie cAMP oder aber meist durch Phosphorylierung regulierte Enzyme sein. Für diesen auch als Signaltransduktion bezeichneten
Vorgang stehen eine Reihe von Mechanismen zur Verfügung, bei denen eine Familie von Guaninnucleotid-bindenden Proteinen , die sogenannten G-Proteine , eine
wichtige Rolle spielt.
G-Proteine kommen in zwei unterschiedlichen Zuständen vor, die sich nur durch das jeweils gebundene Guaninnucleotid unterscheiden. In aktiver Form sind sie mit GTP beladen und imstande, eine
Reihe unterschiedlicher Proteine zu aktivieren. Für die Ueberführung der aktiven in die inaktive Form des G-Proteins ist eine GTPase-Aktivität notwendig, die meist
eine Eigenschaft des G-Proteins selbst ist , häufig aber einen aktivierenden Hilfsfaktor , ein sogenanntes GTPase-aktivierendes Protein (GAP), benötigt.
Soll das inaktive, GDP-beladene G-Protein wieder in die aktive Form überführt werden, so ist zunächst die Abdissoziation des GDP notwendig. Hierfür werden Proteinfaktoren benötigt,
die allgemein als Guaninnucleotid-releasing-Proteine (GNRP) bezeichnet werden. Das Guaninnucleotid-freie G-Protein hat eine hohe Affinität für GTP und
nimmt dies rasch auf, womit es wieder in den aktiven Zustand überführt wird.
Besonders umfangreich ist die Familie der heterotrimeren, grossen G-Proteine. Deren Aktivierung-/Inaktivierungscyclus beginnt mit der Anlagerung der a -Untereinheit an die b g
-Untereinheiten. Gleichzeitig erfolgt die Abdissoziation des gebundenen GDP, wobei ein durch den entsprechenden Liganden aktivierter Rezeptor mit sieben Transmembrandomänen als GNRP dient.
Der Komplex aus aktiviertem Rezeptor sowie dem leeren heterotrimeren G-Protein bindet nun GTP, was zur Abdissoziation des aktivierten Rezeptors führt. Die a -Untereinheit des G-Proteins ist
nun aktiv und assoziiert mit den für die biologische Antwort verantwortlichen Proteinen, die dadurch aktiviert bzw. gelegentlich inaktiviert werden und häufig Reaktionskaskaden auslösen.
Eine intrinsische GTPase-Aktivität der a -Untereinheit sorgt für ihre Inaktivierung und damit für die Signallöschung.
Rezeptoren, die an das Adenylatcyclasesystem gekoppelt sind
Das Adenylatcyclasesystem besteht aus Rezeptoren, heterotrimeren G-Proteinen und Adenylatcyclasen
Eine grosse Zahl von Hormonen bedient sich des Adenylatcyclasesystems zur Signaltransduktion. Das als extrazellulärer Botenstoff dienende Hormon wird auch als erster Informationsträger
oder first messenger bezeichnet. Es bindet an einen in die Plasmamembran integrierten spezifischen Rezeptor. Dies führt über ein heterotrimeres G-Protein zu einer Aktivitätszunahme
der auf der Innenseite der Zellmembran lokalisierten katalytischen Einheit, der Adenylatcyclase. Diese katalysiert die Reaktion:
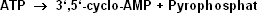
Zunächst gehören zum Adenylcyclasesystem die Rezeptoren für stimulierende bzw. hemmende Hormone oder Substanzen (Rs bzw. Ri). Die Bindung des
entsprechenden Effektors an den Rezeptor wird als Signal auf das entsprechende heterotrimere G-Protein weitergeleitet. Die aktivierten Rezeptoren dienen als GNRPs und lösen die Freisetzung des
gebundenen GDP aus, wodurch es zur Dissoziation der Untereinheiten kommt. Dadurch wird die für den stimulierenden Effekt wichtige Untereinheit Gsα freigesetzt.
Diese bindet ein GTP und ist in dieser Form zur Aktivierung der katalytischen Untereinheit C, der eigentlichen Adenylatcyclase, imstande, was zu einer verminderten cAMP-Bildung führt.
Interessanterweise erfolgt die Informationsübertragung im Fall inhibitorischer Effektoren auf ganz ähnliche Weise . Auch hier löst die Beladung des inhibitorischen
Rezeptors Ri mit seinem Effektor eine Dissoziation eines G-Proteins aus drei Untereinheiten aus. Die inhibitorische a -Untereinheit Gi a ist nach Bindung von GTP imstande, die aktive Form der katalytischen
Untereinheit C in die inaktive zu überführen und so die cAMP-Produktion zu hemmen.
Die stimulierende Untereinheit Gsα verfügt ebenso wie die hemmende Giα über eine intrinsische GTPase-Aktivität. Diese ist für die hydrophobe Spaltung des
gebundenen GTP zu GDP und anorganischem Phosphat verantwortlich, was zur Löschung des stimulierenden bzw. inhibitorischen Signals und darüber hinaus zur Reassoziation der jeweiligen G-Protein-Komplexe
führt. Ein GTPase aktivierendes Protein ist hierfür nicht notwendig.
Es ist von besonderem medizinischem Interesse, dass die Effekte einiger Bakterientoxine offensichtlich über Wechselwirkungen mit den G-Proteinen vermittelt werden .
So führt beispielsweise das Choleratoxin zu einer irreversiblen Aktivierung der Gsα-Untereinheit , was das Adenylatcyclasesystem in einen permanent aktiven Zustand überführt.
Das Toxin des Keuchhustenerregers führt zu einer permanenten Hemmung der Giα -Untereinheit.
Spezielle Funktion der βγ-Untereinheiten der G-Proteine
Es gibt Hinweise für eine besondere Funktion der βγ-Untereinheiten der G-Proteine. Solange die a -Untereinheit GDP gebunden hat, bilden die β- und γ-Untereinheiten mit
ihr ein Heterotrimer. Der Austausch von GDP gegen GTP führt zu einer Dissoziation der a -Untereinheit , die β- und γ-Untereinheiten bleiben jedoch noch relativ fest aneinander gebunden.
In dieser Form sind sie imstande, andere Proteine zu binden. So gibt es Hinweise dafür, dass sie die Phospholipase C und besondere Rezeptorkinasen binden können.
Proteinkinase A durch cAMP aktiviert
Soweit man bis heute weiss, hat cAMP eine einzige intrazelluläre Funktion: cAMP aktiviert eine spezifische Proteinkinase , die auch als Proteinkinase A (PK
A) bezeichnet wird. Es handelt sich um ein tetrameres Enzym, welches aus je zwei unterschiedlichen Untereinheiten, der regulatorischen R-Untereinheit sowie der katalytischen
C-Untereinheit besteht. In Abwesenheit von cAMP werden durch die R-Untereinheiten die Substratbindungsstellen der C-Untereinheiten blockiert. Die Bindung von jeweils zwei cAMP-Molekülen
an jede der zwei R-Untereinheiten führt zu einer Konformationsänderung, die eine Abdissoziation der beiden C-Untereinheiten auslöst, deren Substratbindungsstellen freilegt und sie
somit katalytisch aktiv macht.
Erhöhte zelluläre cAMP-Spiegel beeinflussen nicht nur Stoffwechselenzyme, sondern auch die Transkription spezifischer Gene . Diese enthalten in ihrer Promotorregion
eine Sequenz, die als c-AMP-response-element oder CRE bezeichnet wird. Die Aktivierung von Genen, die CRE als enhancer-Element enthalten, erfolgt nach Bindung
eines Transkriptionsfaktors , des CREB (engl. cAMP response-element binding protein). Das dimere CREB wir durch die zu diesem Zweck in den Kern
translozierte PK A phosphoryliert und kann danach mit einem weiteren Transkriptionsfaktor TF II D sowie der RNA-Polymerase II assoziieren, die TATA-Box (Promotor-Region) lokalisieren und die Transkription
des jeweiligen Gens stimulieren bzw. auslösen.
Rezeptoren, die an die Phospholipase Cβ gekoppelt sind
Für eine Reihe von Hormonen ist gesichert, dass sie nach Wechselwirkung mit ihrem in der Plasmamembran lokalisierten Rezeptor einen Anstieg der intrazellulären Ca2+-Konzentration auslösen,
was zu hormonspezifischen Aenderungen des Stoffwechsels der Zielzellen führt. Das Inositol-(1,4,5)-triphosphat (InsP3) ist der intrazelluläre Ueberträgerstoff, der für
die Erhöhung der Calciumkonzentration veantwortlich ist.
InsP3 entsteht durch Spaltung eines spezifischen Membranphospholipids , des Phosphatidylinositol-4,5-bisphosphat. (PIP2). Dieses wird durch zweimalige, ATP-abhängige
Phosphorylierung von Phosphatidylinositol gebildet. Für die Bildung von InsP3 aus PIP2 wird eine spezifische Phospholipase C, die Phospholipase Cβ benötigt
. Ein weiteres Reaktionsprodukt ausser InsP3 ist Diacylglycerin (DAG), welches in der Membran zurückbleibt.
Für die Erzeugung von InsP3 aus PIP2 sind Rezeptoren mit sieben Transmembrandomänen verantwortlich. Es handelt sich um die a 1-Rezeptoren von Katecholaminen sowie Rezeptoren für
Acetylcholin, Histamin, Angiotensin, Vasopressin, Serotonin und viele Geruchsstoffe. Die Bindung des jeweiligen Hormons (oder Liganden) an seinen Rezeptor führt über ein heterotrimeres
G-Protein nach Austausch von GDP mit GTP zur Aktivierung der Phospholipase Cβ. Interessanterweise binden Tyrosinkinaserezeptoren nach Aktivierung die Phospholipase Cγ , was ebenfalls
zur Spaltung von PIP2 führt .
InsP3 ist imstande, die cytoplasmatische Calciumkonzentration zu erhöhen . Die Calciummobilisierung erfolgt hierbei im wesentlichen aus intrazellulären Calciumspeichern,
welche im endoplasmatischen Reticulum lokalisiert sind. Dort findet sich auch der InsP3-Rezeptor. Insgesamt bilden vier derartige Rezeptormoleküle ein Homotetramer, das einen durch den Liganden
InsP3 aktivierten aktivierten Calciumkanal darstellt.
Jede Erhöhung der cytosolischen Calciumkonzentration führt zu markanten Aenderungen des Zellstoffwechsels. So kommt es u.a.
- Zu einer Stimulierung des Glykogenabbaus in Leber,
Muskulatur und Fettgewebe ,
- Zu einer Stimulierung sekretorischer und kontraktiler Prozesse
- Sowie zur Verstärkung einer Reihe von Effekten, die eigentlich durch cAMP vemittelt sind.
Calcium-bindende Proteine
Calcium wirkt meist nicht direkt, sondern unter Einschaltung calciumbindender Proteine auf die von ihm beeinflussten enzymatischen Systeme ein. In Muskelzellen ist das calciumbindende Protein das Troponin.
In Nichtmuskelzellen wurde als calciumbindendes Protein das Calmodulin nachgewiesen.
Calcium-Calmodulin-abhängige Enzyme sind beispielsweise die Adenylatcyclase oder die membrangebundenen Ca-ATPasen.
Bei der oben geschilderten Spaltung von Phosphatidylinositol-bisphosphat wird nicht nur InsP3 als second messenger zur intrazellulären Calciummobilisierung erzeugt. Auch das frei
werdende Diacylglycerin hat eine wichtige Funktion bei der Signaltransduktion . Es aktiviert zusammen mit Calcium eine spezifische Proteinkinase, die Proteinkinase C.
Rezeptoren mit Tyrosinkinaseaktivität
Bei einer anderen Gruppe von Hormon-Rezeptoren handelt es sich um Polypeptide mit nur einer hydrophoben, die Membran durchspannenden Domäne. Die funktionellen Rezeptoren entstehen nach Bindung
des jeweiligen Liganden und sind Homodimere (Dimerisierung nach Ligandenbindung), im Fall der Rezeptoren für Insulin und IGF-I posttranslational entstandene
Heterotetramere. Allen Rezeptoren ist gemeinsam, dass die Bindung des Hormons eine oder mehrere Autophosphorylierungen an Tyrosylresten des Rezeptorproteins auslöst .
Der Rezeptor ist demnach ein integrales Membranprotein, das über eine ligandenaktivierte Tyrosinkinase verfügt, die eine Autophosphorylierung katalysiert. Dabei werden in der Regel nicht
nur ein, sondern mehrere für den jeweiligen Rezeptor spezifische Tyrosylreste phosphoryliert.
Die phosphorylierten Tyrosylreste dienen als Erkennungssignale für die Anlagerung einer Reihe von Adapterproteinen. Diese verfügen über eine hierfür notwendige Domäne,
die als SH2-Domäne bezeichnet wird.
Ein Enzym mit SH2-Domänen ist die Phospholipase Cγ, die zur InsP3-Bildung führt. Auch das GTPase-aktivierende Protein (GAP)
verfügt über SH2-Domänen und beschleunigt die Ueberführung von kleinen G-Proteinen der Ras-Familie in die inaktive Form. Aktiviert wird Ras durch einen mehrstufigen
Vorgang : das Adapterprotein GRB2 bindet über eine SH2-Domäne an den aktivierten Rezeptor und bindet dann ein als SOS bezeichnetes Protein,
das am in die Membran integrierten Ras-Protein zu einem Austausch von GDP gegen GTP führt. GTP-Ras ist ein Aktivator einer als Raf bezeichneten Proteinkinase, die das erste Glied in einer Kaskade
von Proteinkinasen darstellt. Ihre letztes Ziel sind Transkriptionsfaktoren, welche für die Auslösung von Mitosen verantwortlich sind (MAP Mitogen aktivierte Proteinkinase).
Unterschiedliche Rezeptoren benutzen gleichartige Signalwege
Das Phänomen, dass unterschiedliche Rezeptoren gleichartige Signawege benutzen können, wird im Englischen auch als Receptor Crosstalk bezeichnet. Ein Beispiel hierfür
sind die zwei unterschiedlichen Mechanismen zur Aktivierung des Phosphatidylinositolcyclus. Der eine benutzt G-Protein-verknüpfte Rezeptoren (Phospholipase Cβ),
der andere benutzt Rezeptoren mit Tyrosinkinaseaktivität (Phospholipase Cγ). Beide führen gleichfalls zu einer InsP3-Freisetzung aus PIP2 und damit
zu einer Erhöhung der zellulären Calcium-konzentration.
GH | PRL | Zytokinrezeptoren
Eine andere Familie von Hormonrezeptoren ist diejenige der GH/PRL / Zytokinrezeptoren. Ausser dem Rezeptor für STH (GH = engl. growth hormone) sowie Prolaktin (PRL)
gehören hierzu die Zytokinrezeptoren , speziell die Rezeptoren für die Interleukine und das Erythropoietin.
Die Rezeptoren dieser Familie liegen meist in monomerer Form vor. Die Bindung des entsprechenden Liganden löst eine Dimerisierung des Rezeptors aus und aktiviert gleichzeitig an den Rezeptor
assoziierte tyrosinspezifische Proteinkinasen , die auch als JAK's bezeichnet werden. Aktivierte JAK's phosphorylieren nicht nur sich selbst, sondern auch spezifische Tyrosylreste
der zugehörigen Rezeptoren. Ueber SH2-Domänen binden als STAT's (= engl. signal transducers and activators of transcription) bezeichnete Transkriptionsfaktoren an
diese Tyrosylphosphate (Adapterproteine). Sie werden anschliessend von den assoziierten JAK's phosphoryliert, erlangen dadurch die Fähigkeit zur Dimerisierung und zur Passage in den Zellkern,
wo sie die Transkription spezifischer Gene aktivieren.
Cyclo-GMP als intrazellulärer Botenstoff
In das System von Hormon- und Hormonrezeptorfamilien lässt sich das NO / cGMP-System schlecht einordnen.
Auf hormonelle Signale durch das natriuretische Atriumpeptid , aber auch nach gesteigerter NO-Produktion erfolgt ein Anstieg der intrazellulären Konzentration
von cyclo-GMP (cGMP). Dieser second messenger entspricht in seinem Aufbau dem cAMP, nur dass er aus GTP gebildet wird.
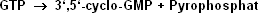
Die durch das natriuretische Atriumpeptid stimulierte membrangebundene Guanylatcyclase unterscheidet sich vom Adenylatcyclasesystem dadurch, dass es sich um ein einziges Transmembranprotein
handelt, auf dessen extrazellulärer Seite die Bindungsstelle für den aktivierenden Liganden liegt (keine vorgeschalteten G-Proteine!).
Ausser der membrangebundenen Guanylatcyclase findet sich in sehr vielen Zellen eine lösliche Guanylatcyclase. Es handelt sich um ein dimeres Enzym aus zwei identischen Untereinheiten,
welches in einem inaktiven und einem aktiven Zustand vorkommt. Das Verhältnis von inaktiver zu aktiver Form des Enzyms hängt von der Konzentration von Nitroxid (NO) ab: je
höher die NO-Konzentration, desto höher die Guanylatcyclaseaktivität .
NO ist ein in der jüngsten Zeit entdecktes intra- und interzelluläres Signalmolekül . Es entsteht enzymatisch aus Arginin.
Im Gegensatz zu cAMP, welches als einzigen intrazellulären Liganden die Proteinkinase A hat, besitzt cGMP eine grössere Zahl von intrazellulären cGMP-Bindungsproteinen.
cGMP-abhängige Proteinkinasen finden sich in besonders hoher Konzentration in glatten Muskelzellen, Thrombozyten und im Kleinhirn. Die wichtigste Funktion der cGMP-abhängigen
Proteinkinase der glatten Muskulatur beruht auf ihrer relaxierenden Wirkung . Dies ist übrigens auch das therapeutische Prinzip aller NO-freisetzenden Vasodilatatoren wie dem
zur Behandlung der coronaren Herzkrankheit eingesetzten Nitroglycerin oder dem zur Behandlung von Erektionsschwierigkeiten eingesetzten Viagra. Man nimmt an, dass sie eine in
der Plasmamembran lokalisierte Calcium-ATPase phosphorlyieren und dadurch stimulieren, so dass die cytosolische Calciumkonzentration in der glatten Muskulzelle absinkt. Auch in Thrombozyten
ergibt sich unter dem Einfluss der aktivierten cGMP-abhängigen Proteinkinase eine Verminderung der zellulären Calciumkonzentration, was den aggregations-hemmenden Effekt von NO erklärt.
cGMP-abhängige Ionenkanäle finden sich in den Photorezeptoren, dem olfaktorischen Epithel sowie dem Epithel der renalen Sammelrohre. In den ersteren der beiden Systeme spielt der cGMP-abhängige
Natriumkanal eine wichtige Rolle bei der Aufrechterhaltung des im unstimulierten Zustand niedrigen Membranpotentials dieser Zelle. Erst durch Aktivierung einer cGMP-spezifischen Phosphodiesterase kommt
es zur Hyperpolarisation dieser Sinneszellen und damit zur Erregungsleitung. In den Sammelrohrepithelien ist der cGMP-abhängige Natriumkanal möglicherweise verantwortlich für die Stimulierung
der Natriurese durch das natriuretische Atriumpeptid. |