Freie Kombination
Freie Kombination findet statt, wenn 2 oder mehr Gene in nicht homologen
Chromosomen lokalisiert sind!!
› siehe Praktikumsaufgabe (2 Genloci betrachtet)
Beispiel Drosophila:

Dihybrider Erbgang
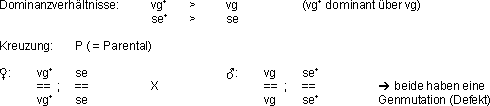
Beide Elternteile sind Homozygoten!
Nachkommen F1:
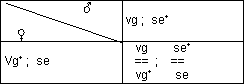
Alle Nachkommen der F1-Generation sind (gezwungenermassen)
Heterozygoten.
Phänotyp der F1-Nachkommen: Wildtyp!
F1 inter se gekreuzt:
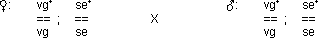
Nachkommen F2:

In der F2 Generation gibt es 4 verschiedene Phänotypen
im Verhältnis 9 : 3 : 3 : 1 (Mendel, 1865) und 9 verschiedene Genotypen!
Wahrscheinlichkeitsverteilung in der F2 bei vollständiger
Dominanz je eines Allels

Beim monohybriden Erbgang ist das Verhältnis der beiden Phänotypen
3 : 1 für das dominante Phän. Beim dihybriden Erbgang ist das
Verhältnis 9 : 3 : 3 : 1.
Testkreuzung für freie Kombination
Man führt eine Rückkreuzung durch: Kreuzung von einem heterozygoten
? mit einem homozygot rezessiven Eltern-?. Falls freie Kombination vorliegt,
kommt es zu folgendem typischen Verlauf:
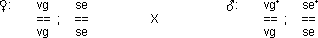
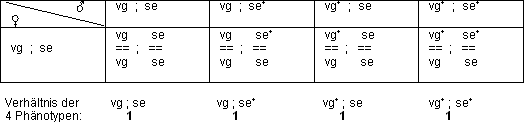
› Beweis für freie Kombination!
Dieses Ergebnis der Kreuzung beweist, dass vg und se auf verschiedenen,
nicht homologen Chromosomen lokalisiert sind.
Einschränkung der freien Kombination durch Kopplung
Wenn zwei Gene auf zwei verschiedenen Chromosomen liegen, ist die freie
Kombination möglich (vg+ auf dem 2. Chromosom, se+
auf dem 3. Chromosom). Die beiden Gene vg+ und pr+
jedoch befinden sich beide auf dem 2. Chromosom (Schreibweise: vg pr). Sie
sind gekoppelt!
Die Kopplung schränkt die freie Kombination ein:
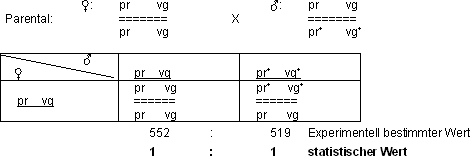
Die zwei Phänotypen stehen im Verhältnis 1 : 1. Das deutet auf
eine absolute Kopplung hin.
› Bemerkung: Bei Drosophila-Männchen gibt es
keine Crossing over (C.O.)!
Schlussfolgerung: › pr kombiniert frei mit se (ist
auch experimentell bestätigt)
Kopplungsregeln:
wenn ein Gen a mit Gen b gekoppelt ist, und wenn Gen b mit Gen c gekoppelt
es, dann ist auch Gen a mit Gen c gekoppelt.
wenn Gen a mit Gen b frei kombiniert, und b mit c gekoppelt ist, so kombiniert
a frei mit c.
Anzahl der Kopplungsgruppen = Anzahl der haploiden Chromosomen (n)
Anzahl Kopplungsgruppen:
Drosophila
Erbse
Triton
Mais
Maus
Mensch |
4
7
12
10
20
23 |
Kopplungsgruppen von Drosophila melanogaster: Auswahl
einiger weniger Gene

Wie schon oben erwähnt, ist die Anzahl der Kopplungsgruppen gleich
der Anzahl der Chromosomenpaare. Bei Dros. mel. existieren 4 Kopplungsgruppen,
sie haben also 4 Chromosomenpaare.
| |
Zahl der bekannten Kopplungsgruppen |
Zahl der Chromosomen (n) |
Drosophila willistoni
Drosophila melanogaster
Drosophila pseudoobscura
Drosophila virilis
Erbse (Gregor Mendel)
Neurospora
Hausmaus
Kaninchen
Mensch |
3
4
5
6
7
7
20
14
23 |
3
4
5
6
7
7
20
22
23 |
Es sind noch nicht ganz alle Kopplungsgruppen bekannt!
Durchbrechung der Kopplung durch Faktorenaustausch als
Folge des „Crossing overs“ in der Meiose
Da bei Dros. mel. nur im Weibchen Crossing over (C.O.) stattfinden, ist
eine einfache Rückverfolgung des Faktorenaustausches gewährleistet.
Kopplung mit Rekombination
reziproke Kreuzung des Tests 1:
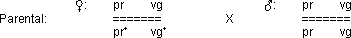
Im Falle einer absoluten Kopplung erhält man zwei Phänotypen.
Hier jedoch erhält man deren vier: es haben sich durch C.O.‘s
Rekombinanten gebildet!

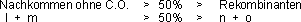
› Es kann sich nicht um eine freie Kombination handeln,
da kein 1 : 1 : 1 : 1 Verhältnis vorliegt.
› Auch eine absolute Kopplung kann ausgeschlossen
werden, da das Verhältnis nicht 1 : 1 ist.
› es handelt sich um eine partielle Kopplung!

1 ME = 1 Morgan-Einheit = 1 % RW
Die Morgan-Einheit ist ein Mass für die Genabstände.
Andere Rekombinationswerte, bzw. Austauschwerte:
vg mit bw (brown)
vg mit L (lobe)
vg mit cn (cinnabar) |
38 % (±)
5 % (±)
10 % (±) |
Diese Werte nennt man biologische Konstante mit Streuung!
diese Werte stimmen nie genau, aber immer ungefähr (d. h. vg mit
cn ist nie 30%, aber vielleicht 7 % oder 12%
das Ergebnis stimmt nur bei einer grossen Anzahl von Versuchen (Gesetz
der grossen Zahlen!)
Zusammenfassung:
Der Testversuch auf Kopplung von zwei Genen besteht aus einem Rückkreuzungstest:
Nachkommen F1 (heterozygotes Männchen mit doppeltrezessiven
Eltern (Weibchen)!
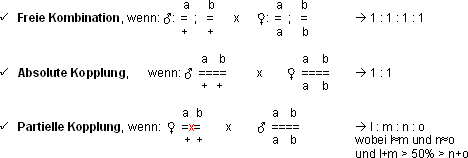
Die lineare Anordnung der Gene
Rekombinationswert - Hypothese:
Der Rekombinationswert (RW) ist ein Mass für die Genabstände
auf den Chromosomen. Die Distanz wir in Morgan-Einheiten angegeben (die
These wurde postuliert von T. H. Morgan und A. H. Sturtevant)
Die RW-Hypothese gilt für sämtliche Lebewesen (Organellen, Zellen,
Bakterien, Pflanzen, Tiere).
Summen-Differenz-Regel:
Genloci lassen sich mathematisch und graphisch bestimmen!
Versuch von Morgan und Sturtevant:

› Zuerst werden Kreuzungen durchgeführt, dann
berechnet man die RW‘s. Durch Addition und Subtraktion können
die Distanzen zwischen den Loci bestimmt werden.

= Summe- oder Differenzregel:
Die Summe- oder Differenzregel beweist die RW - Hypothese, also die
lineare Anordnung der Gene
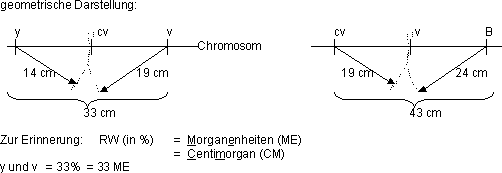
Mit diesem Experiment werden nur die relativen Abstände berechnet!
Die Zuordnung eines Gens an eine bestimmte Stelle des Chromosoms kann nicht
erfolgen! Mindestens einen Genort muss man willkürlich wählen
(meistens den am meisten links liegenden). Beim Chromosom I von Dros. mel.
ist das y (yellow) mit der Position 0.0. Alle anderen Positionen wurden
experimentell bestimmt.
Cytologischer Austausch in der Meiose (Crossing over)
Gesetzmässigkeiten
Beweis, dass die genetischen Rekombinationen auf cytologischem Chromosomenaustausch
(Crossing over) beruhen:
1931 C. Stern: bei Drosophila
1931 Creighton und Mc Clintock bei der Maus
4 Regeln:
Das C.O. geschieht während dem Tetradenstadium (Tetrade, weil 4 Chromosomen;
4=Tetra), d. h. nach vollzogener Paarung der homologen Chromosomen. Tetradenstadium:
von Zygotän bis Pachytän!
Nur zwei von vier Chromatiden sind an einem C.O. beteiligt (Berührungspunkte
= Chiasma).
Freier Wechsel der Stränge für mehrere C.O.‘s pro Tetrade
(alle Stränge sind gleichberechtigt).
Interferenz bei weniger als 10 ME für den Eintritt eines 2. C.O.
Ein C.O. setzt die Wahrscheinlichkeit eines zweiten C.O.‘s in der
Nachbarschaft herab (± regelmässige Anordnung der C.O.). Im
Umkreis von 10 ME von einem C.O. ist die Wahrscheinlichkeit eines 2. C.O.‘s
gleich null.
Rekombination ≤ 50%
Wenn der RW = 50% ist, dann ist das Resultat nicht mehr unterscheidbar
von der freien Kombination, da dann das Verhältnis auch 1 : 1 : 1 :
1 beträgt.

Bei Drosophila melanogaster erlaubt die reziproke Kreuzung eine Unterscheidung
zwischen freier Kombination und Rekombination mit Kopplung! Grund: die Reziprokkreuzung
bei der Rekombination mit Kopplung ergibt ein Verhältnis von 1 : 1
(und nicht 1 : 1 : 1 : 1 wie bei der freien Kombination), weil es beim Männchen
keine C.O. gibt.
2. Der RW ist im Mittel nie › 50%, weil:
Mehrfach - C.O.‘s zwischen zwei weit entfernten Genen vorkommen
nur einfache oder andere ungeradzahlige Austauschvorgänge zwischen
zwei Genen eines Stranges zu Rekombination führen
nur zwei Chromatiden an einer Stelle ein C.O. eingehen können
Einfache C.O. zwischen a und b
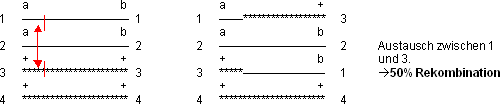
1 und 2 (3 und 4) sind Schwesterchromatide und somit genetisch identisch
2 und 3 (1 und 4) sind homologe Chromatide und genetisch nicht identisch
C.O. zwischen Schwesterchromatiden sind möglich, verlaufen aber unbemerkt
Weitere Austauschmöglichkeiten: 1/4, 2/3, 2/4. Sie führen jeweils
zu max. 50% Rekombination.
Bei einem C.O. hat man maximal 50% Austausch!
Doppeltes C.O. zwischen a und b
Fall 1:
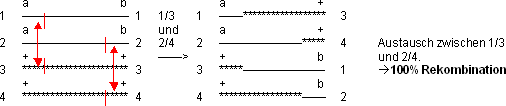
Weitere Austauschmöglichkeiten: 1/4 und 2/3; 2/3 und 1/4; 2/4 und
1/3
Fall 2:
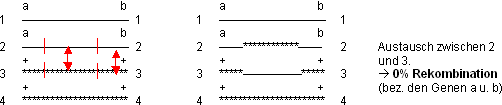
Weitere Austauschmöglichkeiten: 1/3 und 1/3; 1/4 und 1/4; 2/4 und
2/4
Bemerkung: Wie betrachten hier nur die Gene a und b. Die C.O.‘s, welche
die Gene auf den mittleren Segmenten betreffen, können nur mit sehr
präzisen und aufwendigen Tests nachgewiesen werden.
Fall 3:
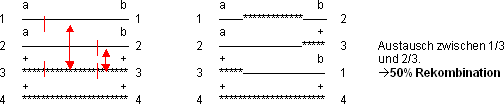
Weitere Austauschmöglichkeiten: 1/3 und 1/4; 1/4 und 1/3; 1/4 und
2/4; 2/3 und 2/4; 2/4 und 1/3; 2/4 und 1/4; 2/4 und 2/3

Die Fälle 1 und 2 treten statistisch mit der gleichen Frequenz auf
(haben also zusammen eine Rekombination von 50%), der Fall 3 führt
zu 50% Austausch:
› statistisch 50 % Rekombination bei 2 C.O.‘s!
Merke: Der RW überschreitet auch für n = 3,4,5,6,
... n C.O.‘s die 50% nicht (statistisch).
Genkarten
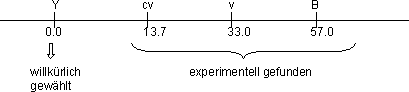
Konstruktion von Genkarten: y = links, an der Position 0.0. Diese Position
wird willkürlich gewählt. Hätte man weiter links noch ein
Gen gefunden, wäre nicht y an der Position 0.0! Alle anderen Positionen
der Gene werden berechnet und addiert.
Beziehung zwischen RW und Genkarte:
für kleine Distanzen ≤ 10 ME: Mapdistance = RW,
da die Interferenz ein zweites C.O. hemmt.
für grössere Distanzen › 10 ME: Mapdistance ›
RW, da beim Auszählen nicht alle Rekombinanten erfasst werden
(Doppel-CO’s).
Aufbau der Genkarte: Der Aufbau der Karte erfolgt durch die Ermittlung
kleinerer Abstände, die addiert werden! Bei kleinen Abständen
werden Doppel-CO’s durch Interferenz verunmöglicht.
Spezialbefunde im heterogametischen Geschlecht (XY):
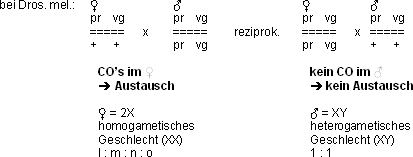
Im heterogametischen Geschlecht (XY) kein CO, also kein Austausch:
Dros. Männchen
Seidenspinner Weibchen (XY, es gibt Tiere, da ist das Weibchen heterogametisch,
u.a. auch Tauben)
Warum es im heterogametischen Geschlecht keinen Austausch gibt, weiss
man auch heute nach 90 Jahren Drosophila-Forschung noch nicht.
Im heterogametischen Geschlecht verminderter Austausch:
Im Männchen weniger Austausch als im Weibchen: Maus, Ratte, Mensch
Im Weibchen weniger Austausch als im Männchen: Haustaube
Im Weibchen gleich viel Austausch wie im Männchen: Erbse
Wenn die Häufigkeit der C. O. vermindert ist, sind auch die berechneten
Abstände auf der Genkarte kleiner. In Wirklichkeit aber, auf der
cytologischen Karte also, sind sie gleich gross bei Weibchen und Männchen!
Genetischer Abstand ≠ cytologischer Austausch
Frage: Kann man eine Genkarte eines Dros-Männchen machen?
› Nein, man kann nur eine des Weibchens machen und
diese dann auf das Männchen transferieren.
Polytäne Chromosomen und Genkartierung durch Deletion
Polytän = vielfädig, vielstränig
Polytäne Chromosomen = vielstränige Riesenchromosomen, bestehend
aus vielen Chromatiden, mikroskopisch sichtbar
Entdeckung und Vorkommen
Entdeckung:
Heitz und Bauer: 1933 entdeckt bei Mücke (Bibio hortulanus)
Painter: 1933 entdeckt bei Dros. mel.
Vorkommen:
Polytäne Riesenchromosomen kommen hauptsächlich in bestimmten
Insektenorganen (Dipteren-Larven) vor, nicht aber in Muskel- und Nervengewebe.
In Larven:
- Speicheldrüsen
- Malpighsches Gefäss
- Darmzellen
- Fettkörper
In Adulttieren:
- Nährzellen der Ovarien
- Borstenbildende Zellen
Die Riesenchromosomen entstehen in Zelltypen, die sich selbst nicht mehr
teilen, deren Chromosomen sich aber noch verdoppeln. Die polytänen
Chromosomen organisieren sich zu charakteristischen Fibrillenbündeln
(aus bis zu tausend Einzelfäden).
Jedes Chromosom besteht aus einem charakteristischen Muster: Scheibchen
entsprechen Eu- und Heterochromatin (unterscheiden sich in der Menge oder
Konzentration der DNA). Das Euchromatin (verliert im Ruhekern seine Färbbarkeit)
enthält mehr Gene als das Heterochromatin.
Feststellung: Bandzahl ≈ Genzahl! Das legt die Vermutung nahe, dass
die Bänder den Genen entsprechen. Es wurde aber gezeigt, dass Gene
über mehrere Bänder gehen und ein Band mehrere Gene enthalten
kann.
Die am klarsten strukturierten Riesenchromosomen findet man in den Speicheldrüsenzellen:
Enge Paarung der Homologen
Auch in den Interphasen sichtbar
Heterochromatin (spezifisch färbbar und so unterscheidbar vom Euchromatin)
ist um die Centromerregion (Chromozentrum) konzentriert.
Bau der polytänen Chromosomen und Vergleich mit gewöhnlichen
Chromosomen
Grundsätzlich: Euchromatin anders färbbar als Heterochromatin
(früheres Unterscheidungskriterium).
Im Euchromatin sind fast alle Gene enthalten, während das Heterochromatin
beinahe genleer ist. Das Heterochromatin geht später in die Teilungsphase
(S-Phase) als das Euchromatin.
Was ist bei Riesenchromosomen anders?
Sie sind ca. 300 mal länger (durch maximale Streckung des DNA-Fadens
zwischen den Banden)
Polytäne Chromosomen: vollständig gepaart
Gewöhnliche Chromosomen: › distance gepaart
Sie sind viel dicker. Dieser riesige Durchmesser entsteht durch wiederholte
Replikation der Chromatiden, ohne dass sie sich anschliessend trennen (210
= 1024 Replikationen). Die 1024 Chromatiden bilden ein Band auf dem Riesenchromosom.
Im Normalfall gilt: 1 Bande = 1 Gen
Es kommt aber auch vor, dass eine Bande verschiedene Gene enthält oder
dass sich ein Gen in mehreren Banden befindet!
Heterochromatin: fast genleer; Euchromatin: fast alle Gene enthaltend.
Chromomeren sind Subeinheiten der Banden.
Spezialität bei den Dipteren: somatische Distanzpaarung der Homologen
in den Speicheldrüsenzellen (auch in der Metaphase der Mitose sind
die homologen Chromosomen beieinander - zwar mit bestimmtem Abstand
- was eigentlich in der Mitose nicht nötig wäre). In den
Speicheldrüsenzellen findet man die am klarsten strukturierten Riesenchromosomen.
Die homologen Chromatiden sind gebündelt und bilden ein vielsträniges
Riesenchromosom. Bis zu 1024 Chromatiden (210) können darin enthalten
sein (erklärt die Dicke).
Bedeutung der polytänen Chromosomen
1. Adäquates Substrat für die Theorie der linearen Genanordnung
Bei Drosophila:
ca. 5‘000 Bänder
ca. 5'000 Gene (nicht ganz sicher)
Dies legt die These nahe, dass ein Band einem Gen entspricht. Heute steht
fest, dass diese Annahme zu einfach ist. Ein Gen kann mehrere Bänder
umfassen (z. B. Bar) und es können mehrere Gene auf einem Band liegen.
2. Nachweis von Strukturmutationen
Paarung der Homologen: Es werden genaugenommen die homologen Loci und nicht
einfach das ganze Chromosom gepaart.
Mutation: Deletion! ( = Stückausfall)
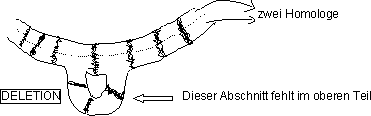
3. Ermöglichen Analyse der Gewebe- und Stadienspezifischen
Genaktivität ( = Genexpression)
Puff: lokale Anschwellung einer Chromosomenregion und
Auflockerung des Bandenmusters. Dies ist der Ort der Genaktivität.
Das Gen, welches angeschwollen ist, ist aktiv.
4. Ermöglichen genaue Kartierung von Genen auf den Chromosomen
Genkartierung durch Deletion
Methode:
Herstellung von Deletionen (= Erzeugung von Stückausfällen durch
Röntgenbestrahlung auf männliche Chromosomen)
Kreuzung dieses männlichen Chromosoms mit rezessivem Weibchen
Konkretes Beispiel:
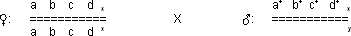
Alle Weibchen Nachkommen der F1 Generation sind
phänotypisch wild, weil die Mutation rezessiv ist:
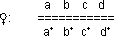
Nun kreuzt man das gleiche Weibchen mit einem Männchen, bei welchem
im X-Chromosom eine Deletion auftritt (z. B. b+ fehlt). Diese Deletion kann
künstlich induziert werden durch Röntgenstrahlen. Die Zerstörung
tritt allerdings zufällig auf, man kann sie nicht steuern (es ist nicht
möglich, gezielt ein Gen ausfallen zu lassen). Die Röntgenstrahlen
führen zu Chromosomenbrüchen (sind also mutagene Agenzien).
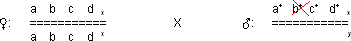
Die Töchter sind nun phänotypisch b, weil das b+ vom Vater fehlt:

Das rezessive b Allel hat sich manifestiert: Pseudodominanz!
Das an sich rezessive Gen b wird scheinbar dominant, in Wirklichkeit aber
bleibt es rezessiv.
Der Ort der Deletion lässt sich nachweisen (cytologische Untersuchung
der Nachkommen). Besonders gut eignen sich Riesenchromosomen für die
cytologische Analyse.
› Der Loci des Gens b+ befindet sich in der Deletion.
Diese Methode findet Anwendung bei der Herstellung von Genkarten (= Genkartierung
mit überlappender Deletion).
Der Nachteil dieser Methode: Deletionen können sehr lang sein, so
dass ganze Regionen ausfallen und nicht nur ein einzelnes Gen. Dies wird
korrigiert, indem die Kartierung mit überlappenden Regionen vorgenommen
wird. Überlappende Deletionen erlauben ein genaues lokalisieren der
Gene.
Vergleich zwischen genetischer und cytologischer Karte
Die Genreihenfolge ist auf beiden Karten identisch.
Die Genabstände allerdings weisen auf der genetischen und der cytologischen
Karte systematische Abstände auf (siehe Beilage 5/10).
Welches ist nun die genaue Karte, die cytologische oder die genetische?
Die cytologische Karte ist die genaue. Die genetische Karte stimmt nicht,
weil die C.O. längs der Chromosomen nicht überall dieselbe Wahrscheinlichkeit
haben. Das führt zu einer Verzerrung der genetischen Karte, welche
auf der Häufigkeit der C.O. beruht.
Genetische Karte: Im Allgemeinen ist die Häufigkeit
von C.O. in der Telomerregion (Chromosomenende) und in der Centromerregion
kleiner als in den übrigen Chromosomenregionen. Das führt dazu,
dass die Gene in der Telomer- und der Centromerregion näher zusammenliegen
auf der gen. Karte.
Ausnahme: Mensch, C. elegans. Hier ist die C.O.-Häufigkeit in der Telomer-
und Centromerregion stark erhöht.
Genetische Karte: entspricht der relativen Lage der Gene aufgrund der C.O.
Cytologische Karte: entspricht der reellen Lage der Gene |
